 Blog
Blog
メダカ個体解析の論文初上梓ーMBoCの表紙を飾る
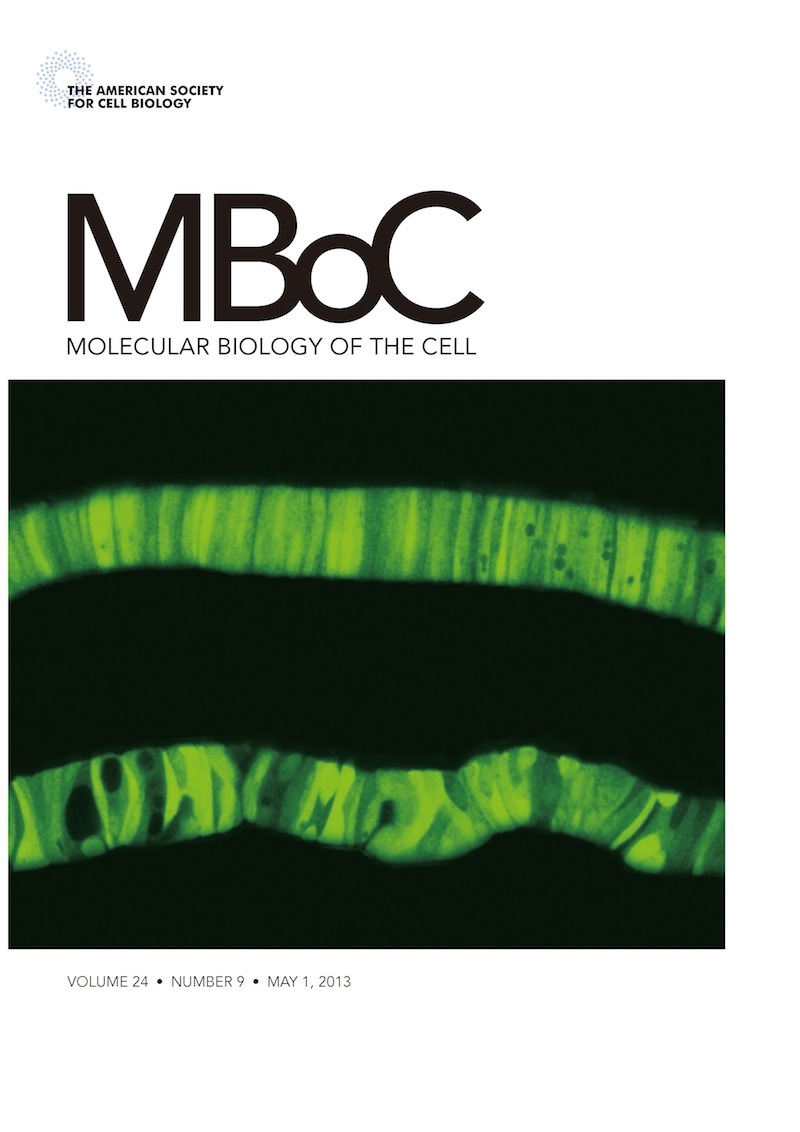 石川助教が6年の歳月を費やして完成させた、初のメダカ個体解析の論文が、米国細胞生物学会が発行する雑誌Molecular Biology of the Cell (MBoC) の5月1日号に掲載されました。しかもHighlight に選ばれ、右図のように表紙を飾りました (ホームページの不具合により、本日5月13日にアップしました)。
石川助教が6年の歳月を費やして完成させた、初のメダカ個体解析の論文が、米国細胞生物学会が発行する雑誌Molecular Biology of the Cell (MBoC) の5月1日号に掲載されました。しかもHighlight に選ばれ、右図のように表紙を飾りました (ホームページの不具合により、本日5月13日にアップしました)。
哺乳動物の小胞体ストレス応答では、IRE1・PERK・ATF6の3経路が機能していて、IRE1経路下流の転写因子XBP1のノックアウトマウスは肝臓の発達不全によって胚生致死となり、PERKのノックアウトマウスは生後糖尿病を発症しますので、小胞体ストレス応答が生理的に重要であることがわかります。ATF6経路(ATF6αとATF6βの2種類が存在)については我々自身でノックアウトマウスを作出し(とても大変でしたが)、ATF6αが小胞体シャペロンの転写誘導に必要であることを示しました。さらに、ATF6α・βそれぞれの単独ノックアウトマウスは表現型を示しませんでしたが、ATF6α・βダブルノックアウトマウスは胚生致死になりました(ATF6βがATF6αのバックアップの役割を果たしていると考えています)。ところが、E8.5という段階でお腹を開けても、ダブルノックアウトマウスが見当たらないので、3経路の中でATF6経路の破壊が最も重篤な表現型を示すことがわかりましたが、E8.5でいないと、死ぬのが早すぎて、我々の手では何が原因で死ぬのか究明することが困難でした。
そこで、母親の胎内で育つマウスと異なり、卵から孵化するメダカを使えば、初期発生過程を顕微鏡下で観察できるので、致死の原因がわかるのではないかと思って、メダカの解析を6年前に石川助教がM1だったときに開始しました。調べて見ると、期待通り、メダカでも上記3経路が機能しており(Ishikawa et al., CSF, 2011)、マウスの場合と同様にATF6α・βそれぞれの単独ノックアウトメダカは表現型を示しませんでしたが、ATF6α・βダブルノックアウトメダカは胚生致死になりました。
致死の原因を調べてわかったのは、ATF6α・βダブルノックアウトメダカでは脊索が発達しなかったのです。脊索は背骨ができる前に体の軸として働く重要な器官です。表紙を飾った写真の上は野生型の脊索で、円盤状の脊索鞘細胞が直線的にきれいに並んでいます。脊索鞘細胞は8型コラーゲンなどの細胞外マトリックスタンパク質を大量に合成分泌していて、細胞の外から自身を支えさせています。このとき、ATF6α・βが活性化され小胞体シャペロンが転写誘導されて、これらのタンパク質の品質が管理されているのです。一方下のATF6α・βダブルノックアウトメダカでは、小胞体シャペロンを転写誘導することができないために細胞外マトリックスタンパク質を十分に分泌することができなくなり、脊索鞘細胞のならびがガタガタになって(所々穴が見えますよね)、脊索の発達が止まることがわかりました。
実際に脊索が発達する段階では、生理的に小胞体ストレスが発生していて、モルフォリノを用いて8型コラーゲンをノックダウンすると、この小胞体ストレスが減じることがわかりました。つまり、8型コラーゲンを大量合成することが小胞体ストレスの原因であることを示すことができました。生理的小胞体ストレスの原因タンパク質を初めて同定することができたと自負しています。
表紙の写真は、小胞体シャペロンBiPのプロモーターを使って発現させたEGFPを観察しています。当然ながら、下のATF6α・βダブルノックアウトメダカではBiPプロモーターが活性化されていませんから、EGFPの発現量は低いです。脊索の形状比較のため、下は上に比べて20倍長い時間観察しています。
ツニカマイシンやタプシガルジンといった薬物を細胞に投与すると、強力に小胞体ストレスを起こすことができますので、小胞体ストレスの分野では、これらを安易に使った非生理的状況下の実験がまかり通っています。これらの薬物は小胞体ストレス応答3経路で働く分子の同定やシグナル伝達機構の解明においては必須の役割を果たしてきましたが、その体勢が判明した今、今後は薬物に頼らない、生理的もしくは病理的小胞体ストレスの研究を行って行かなければならないと思っています。目指せ、薬物依存からの脱却。
ブログの記事
- 奈良、富山、釜山 (初)、幕張
- 研究室旅行 & 中野先生 & APPW2025
- 田中啓二先生追悼行事
- 謹賀新年2025 & 平安神宮奉納
- 少年剣道 洛西大会
- 臨床ストレス応答学会@宮崎
- 青陵高校生の訪問
- インターハイ & 少年剣道全国大会
- 初武漢 & 受賞リストを更新:Frontiers of Knowledge Award
- 今年は中学生チームが全国大会出場!
- 名誉教授 & 初上海
- 最終講義 & 祝賀会 & B B Q
- 謹賀新年2024
- 少年剣道・洛西大会
- 青陵高校生の訪問
- Mary-Jane の来日
- 少年剣道全国大会
- 悲願の全国大会出場 & 近畿支部例会
- 細胞の中の分子生物学 & 薬学会@札幌
- 謹賀新年 & 陳君の結婚式@台湾 & 平安神宮 & 洛西大会
- 九份観光編2
- 九份観光編
- 久留米市で講演 & 青陵生の訪問
- 3年ぶりの海外出張@モントリオール
- 3年ぶりの海外出張@モントリオール - part2
- 謹賀新年2022
- 謹賀新年2021
- 蜷川暁特定助教が生化学会奨励賞を受賞
- 求む森研23期生
- 謹賀新年、インフルエンザ、ラスト・トロント
- 第13回 研究室旅行、その他
- 初のスコットランド、グラスゴー
- 初のスコットランド、インバネス
- 初のスコットランド、エディンバラ
- 倉敷青陵高校生の訪問
- 受賞リストを更新しました(安藤百福賞)。ご覧あれ。
- 京大の東南アジアネットワークフォーラム第13回@マニラ
- 謹賀新年
- 受賞リストを更新しました(文化功労者選出)。ご覧あれ。
- 第12回 研究室旅行
- 長沙、今年2回目の中国
- いつまで続く、平成末の大災害?
- 『細胞の中の分子生物学』2万部越え! 森研21期生募集中
- 求む、森研21期生
- 8年ぶりの北京、4月から生物科学専攻長再び
- 地元の方々の研究室訪問
- 「細胞の中の分子生物学」第7刷
- 「細胞の中の分子生物学」第6刷と特別講座開設
- 受賞リストを更新しました。ご覧あれ。
- 第11回 研究室旅行
- eLifeにメダカ個体解析論文を上梓 & パリ
- 「細胞の中の分子生物学」第5刷と新学術領域研究
- メダカ BBF2H7 解析結果を JCB に上梓
- 求む、森研20期生
- 長崎、薬理学会、龍馬通り
- 大隅先生 ノーベル生理学・医学賞受賞祝賀会
- 初アイルランド、ゴールウェイ
- 初アイルランド、モハーの断崖
- 初アイルランド、ダブリン
- 「細胞の中の分子生物学」第4刷重版なる!
- 第10回 研究室旅行
- 朝日新聞に「細胞の中の分子生物学」の書評掲載
- 少年剣道2016
- 「細胞の中の分子生物学」第3刷重版なる!
- 「院生募集」欄に、蜷川研究員の Nobel Forum 参加感想文を掲載しました。
- ノーベル・フォーラム@カロリンスカ研究所に出席
- ノーベル・フォーラム@カロリンスカ研究所に出席−2
- 大隅良典先生、ノーベル賞のご受賞誠におめでとうございます。
- 「細胞の中の分子生物学」重版出来なる!
- 受賞リストを更新しました (恩賜賞・日本学士院賞)。
- 受賞リストを更新しました (トムソンロイター)。ご覧あれ。
- 一般向け生命科学入門書「細胞の中の分子生物学」を上梓
- インド、プネーの石窟、続き(2つ下、5月25日のブログから読んでください)
- インド・プネーの石窟
- 2回目のインド、ニューデリー&プネー
- 院生募集欄の更新
- 初インド、酵母生物学国際学会@コルカタ(旧カルカッタ)
- 第9回研究室旅行
- EDEM1/2/3 トリプルノックアウト細胞の解析結果を JCB report にて発表
- Cell Stress Society International in China - real part2
- Cell Stress Society International in China - part2
- Cell Stress Society International in China
- FASEB Summer Research Conference 2015
- 母校での講演と倉敷青陵高校生のラボ訪問
- 求む森研18期生
- 受賞リストの更新
- 第8回研究室旅行
- EDEM1/2/3 ノックアウト細胞の解析結果を JCB report にて発表
- 森研発足15周年記念パーティ
- 分子遺伝学シンポジウム 2014
- 受賞リストを更新しました
- 謹賀新年
- 9月23日、下村先生ご来阪
- 9期生石川助教の結婚と5期生安達君の来訪
- メダカ個体解析の論文初上梓ーMBoCの表紙を飾る
- 大学院入試説明会ー求む森研16期生!
- CREST シンポジウム
- 少年剣道
- 第6回研究室旅行
- IUBMB & FEBS at Sevilla, Spain に出席
ブログのカレンダー
 |
2025年7月 |  |
||||
|---|---|---|---|---|---|---|
| S | M | T | W | T | F | S |
| 29 | 30 | 1 | 2 | 3 | 4 | 5 |
| 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 13 | 14 | 15 | 16 | 17 | 18 | 19 |
| 20 | 21 | 22 | 23 | 24 | 25 | 26 |
| 27 | 28 | 29 | 30 | 31 | 1 | 2 |
アーカイブ
- 2025年06月
- 2025年04月
- 2025年02月
- 2025年01月
- 2024年12月
- 2024年11月
- 2024年09月
- 2024年08月
- 2024年07月
- 2024年05月
- 2024年04月
- 2024年03月
- 2024年01月
- 2023年10月
- 2023年09月
- 2023年08月
- 2023年07月
- 2023年06月
- 2023年04月
- 2023年01月
- 2022年09月
- 2022年08月
- 2022年01月
- 2021年01月
- 2020年09月
- 2020年03月
- 2020年02月
- 2019年12月
- 2019年07月
- 2019年06月
- 2019年03月
- 2019年02月
- 2019年01月
- 2018年12月
- 2018年10月
- 2018年05月
- 2018年04月
- 2018年03月
- 2018年02月
- 2017年12月
- 2017年10月
- 2017年08月
- 2017年06月
- 2017年03月
- 2017年02月
- 2016年11月
- 2016年10月
- 2016年09月
- 2016年08月
- 2016年06月
 電話:075-753-4067
電話:075-753-4067 Fax:075-753-3718
Fax:075-753-3718